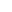【本期导读】
随着组织透明化技术的快速发展,越来越多的生物学应用需要对大规模透明样本进行整体成像,并已在毒理学、神经科学和发育生物学中获得了成功应用。尽管光片荧光显微镜(LSFM)在这一领域得到了广泛应用,但光学投影断层成像(OPT)仍作为一种经典的低成本成像技术,尤其适用于厘米级样本,如透明化的器官、活体胚胎以及植物。OPT的基本原理与计算机断层成像非常相似,涉及通过从不同方向将三维体积投影到二维平面上来捕捉光强分布,并随后通过滤波反投影重建三维结构。然而,传统方法通常由于缺失锥问题和空间非均匀光学像差而面临分辨率下降的问题。此外,在高分辨率成像过程中,通常需要在小景深下进行密集的轴向扫描。
近日,来自清华大学的研究人员提出了一种提出了一种紧凑且低成本的方法,称为扫描光场层析成像(SLiT)系统,用于对大规模样本进行高速高分辨率的整体三维成像,且具有接近各向同性的分辨率。相较于传统OPT则需要数百个投影,SLiT的重建过程仅需要24个投影。同时该系统无需额外的轴向扫描提高景深,进而提高了成像速度。基于先前的数字自适应光学技术,引入了多共轭数字自适应光学(MDAO)修正了多角度间复杂光学像差。整个6 × 6 × 6 mm³的体积可以在30秒内完成采集,并拥有近乎各向同性的高分辨率三维重建(x、z约为17 µm,y约为10 µm)。并在多种样本上展示了SLiT的多功能性和优势,如斑马鱼血管、透明化小鼠大脑、血管染色的透明化小鼠眼睛以及骨染色的透明化小鼠头部。该研究于2024年10月作为封面文章发表于《OPTICA》。

图1 该研究作为封面文章于2024年10月发表于《OPTICA》
【技术背景】
生物成像技术是现代生命科学研究的重要工具,能够帮助科学家观察细胞和组织在生理及病理状态下的动态变化。传统的生物成像方法,如LSFM和OPT,在成像速度和精度上遇到了瓶颈。LSFM作为逐层扫描成像方法,速度存在天然上限。OPT为了满足高分辨率的需求,校准和重建技术不断得到改进,并逐步引入了更加精确的模型。然而,由于非均质样本和变化的成像条件导致的复杂光学像差问题,仍然对高分辨率重建产生不利影响,且仍未得到解决。同时OPT受限于单视图成像的景深,在需要对大面积进行高分辨率成像的场景中,通常需要对大规模样本进行轴向扫描。目前尚无任何现有的OPT系统能够实现对大体积样本的高速整体成像。
光场技术作为一种先进的成像方法,捕捉场景中光线的空间和角度信息,实现了三维场景的感知与重建。传统的成像技术通常仅记录光线在二维平面上的强度信息,而光场技术则进一步记录了光线的传播方向和角度,从而能够在后期处理时生成多视角和深度信息。但是由于像素大小与微透镜大小的限制,光场显微镜的空间分辨率相对较低。为了解决这一问题,扫描光场显微镜使用移动微透镜的方式,提高了空间采样率,进而提高了空间分辨率。然而,由于样本的非均质性和复杂光学环境,光场技术仍然面临视角不一致和像差校正的挑战。为了克服这些问题,当前的光场重建算法逐步引入了更精确的光学模型和多视点融合方法,以提高重建精度和成像速度,从而为大规模三维成像和虚拟现实等应用领域提供更可靠的技术支持。
【技术路线】
SLiT系统中检测组件的机械设计基于水平扫描光场显微镜,通过可旋转的样品架实现多视角数据采集(见图2 (a))。通过简单地更换物镜,分辨率和体积大小可调,以满足样品的需求。在MDAO模型中,光学像差被建模为由不同深度的多个虚拟相位板引起的三维变形。通过在无像差的光学系统模拟中加入虚拟相位板,可以直观地展示像差对成像结果的影响(见图2(b))。为了通过多相位平面像差校正恢复各向同性的高分辨率(见图2(d)),研究人员开发了一个多视角融合和反卷积处理流程(IMPD)。IMPD基于DAOSLIMIT的相位空间迭代反卷积方法,包含相位空间和反投影步骤(见图2(e))。相位空间投影步骤是将估计的体积投影到光场测量值上,而反投影步骤则利用测量值和前向投影之间的误差来更新三维体积。研究人员采用了乘子交替方向法(ADMM)算法,用于体积和相位的迭代同时更新。

图2. SLiT的原理与性能。(a) SLiT与OPT的光学系统比较。与OPT的标准宽场检测臂相比,SLiT的检测臂上的扫描光场设置可以大大扩展景深。SLiT在多视角成像过程中能够覆盖更大的体积成像范围。(b) 复杂成像环境引起的多层相位示意图。当光线穿过非均匀界面时,多次反射会导致成像结果中的像差。(c) 不同平面上的相位会引起不同的像差。当在瞳孔平面和焦平面上没有相位时,光线直线通过,不会对体积造成位移或倾斜;当瞳孔平面上出现相位时,光线会发生位移和倾斜。然而,当相同相位出现在焦平面时,光线仅发生倾斜。这个理想化的例子表明,三维相位引起的体积位移和倾斜是非耦合的,不能仅通过一个平面上的相位来表示。(d) 各个视角引起的像差导致视角之间的信息不匹配,破坏了重建效果。通过应用MDAO,所有像差得到了校正,最大化了重建质量的恢复。(e) ADMM优化策略的结构。重建过程通过IMPD完成。MDAO估计过程通过每个视角的像差模拟过程的迭代反向传播来实现。

图3 (a) MDAO增强了SLiT成像中荧光微球的重建性能及其对应的分辨率(FWHM)。(b) 沿z轴和y轴的最大强度投影(MIP)显示使用SLiT(包含MDAO校正,青色)和不包含MDAO校正(洋红色)的0.055 NA物镜下重建的荧光微球结果。SLiT在整个体积范围内改进了x、y和z轴的分辨率。(c)、(d)、(e)、(f)为从(a)和(b)中白色框标记的区域放大后的MIP图像;从(a)中选取的两个区域的尺寸均为0.5 mm × 0.5 mm × 0.5 mm。(g) 显示SLiT(包含MDAO校正)沿y轴和z轴的横向和轴向平均点FWHM(在(a)的坐标系统中)。整个体积的分辨率几乎是均匀的。由于像差的存在,从不同视角拍摄的图像在x和z方向上的配准精度有限,这导致x和z方向的分辨率(大约17 µm)比y方向的分辨率(大约10 µm)要差。(h) 显示来自24个视角的MDAO相位结果。PP代表瞳孔平面,相FP代表焦平面。(a)和(b)中的比例尺为200 µm。

图4 对比SLiT和LSCM成像的斑马鱼血管结果。(a) 多模态斑马鱼成像结果的三维渲染体积,侧面标记了旋转轴。SLiT的结果显示为黄色,LSCM的结果显示为洋红色。LSCM数据来自视角0。(b) 从视角0、视角45和视角90投影的多模态斑马鱼成像结果的最大强度投影(MIP)。(c) 多模态斑马鱼成像结果的切片堆叠。切片堆叠垂直于视角0进行,在(a)中用白色框标记了堆叠位置。(d)、(e) 为斑马鱼成像结果的子体积MIP图像。SLiT和LSCM的结果分别用黄色和洋红色通道分开。在(a)中用绿色和蓝色框标记了体积位置。(b)、(c)中的比例尺为100 µm,(d)、(e)中的比例尺为50 µm。

图5中尺度透明组织体积成像(a) 小鼠大脑皮层第四层的MIP图:展示了使用SLiT技术,在2倍物镜(数值孔径NA=0.055)下,通过24个视角和单个视角重建的透明小鼠大脑皮层第四层。右下角的图像是3D渲染的透明小鼠大脑皮层第四层。 (b) 神经元的子体积MIP图:从x、y、z三个维度投影的神经元的子体积MIP图。 (c) 神经元强度剖面图:对应于(b)图中的红线。 (d) 透明小鼠眼睛中的血管体积阴影图。 (e) 眼睛的MIP图:从x和z维度投影的眼睛的MIP图。为了显示细节,仅包括一半的眼睛。 (f) 虹膜血管的子体积MIP图:对应于(d)图中指示的视图的子体积。
为了评估SLiT的性能,研究人员使用0.055 NA的物镜对随机分散在1%琼脂糖凝胶中的直径为1 µm的荧光微球进行三维成像。未经MDAO校正时,多视角之间存在严重的错位,出现明显的伪影且分辨率较低。经过MDAO校正后,多视角数据可以被正确对齐,获得更高的空间分辨率且没有伪影(图3(a)–3(f))。此外,通过计算SLiT重建中微球的半高全宽(FWHM),发现实验中横向和轴向分辨率几乎一致(x, z方向约为17 µm,y方向约为10 µm),并且在整个体积范围内经过多位置三维像差校正后保持一致。不同视角的估计相位显示在图3(h)中。视角之间的对齐和与重建结果的相关性验证了MDAO估计的有效性和合理性。
为了进一步评估SLiT在生物样品上的性能,研究人员使用5×/0.14 NA物镜对发育中的斑马鱼血管进行了大体积成像。作为参考,同一样品也使用激光扫描共聚焦显微镜(LSCM)和4×/0.13 NA物镜进行了成像比较。尽管LSCM的结果验证了SLiT的准确性,但由于样品散射的影响,LSCM未能成像斑马鱼背面的眼睛。SLiT以更快的成像速度实现了比LSCM更高的轴向分辨率,这得益于其旋转过程。各种视角的最大强度投影(MIP)显示了血管结构的细节基本一致(图4(b))。某些不匹配可能是由于LSCM的像差以及样品存储期间斑马鱼的生长引起的。在视角90下,LSCM图像在斑马鱼背面因前部组织散射而变淡。生成的切片堆叠展示了血管结构的细节及断层成像的能力(图4(c))。几个子体积被进一步放大,以显示SLiT的分辨率提升(图4(d)和4(e))。在视角0下,SLiT(黄色)与LSCM的血管结构大体一致。然而,在45和90视角下,LSCM的结构由于较低的轴向分辨率而变得模糊,而SLiT仍能显示出清晰的结构。
为了进一步展示SLiT在大尺度透明化组织上的优势,研究人员在多种不同的尺度下对几种不同类型的样品进行了成像,包括透明化小鼠大脑中的神经元(图5)、透明化小鼠眼睛中的血管(图5)以及透明化的染骨小鼠头部。
 邮箱:qhdai@tsinghua.edu.cn
邮箱:qhdai@tsinghua.edu.cn 地址:清华大学中央主楼三楼
地址:清华大学中央主楼三楼 传真:+86 10 62773433
传真:+86 10 62773433